News


24 June 2024
Stowers Faculty EDGE Workshop prepares postdocs for the academic job market
The Faculty EDGE series is just one of many advantages of the postdoctoral training program at the Stowers Institute.
Read Article
By Elise Lamar, PhD
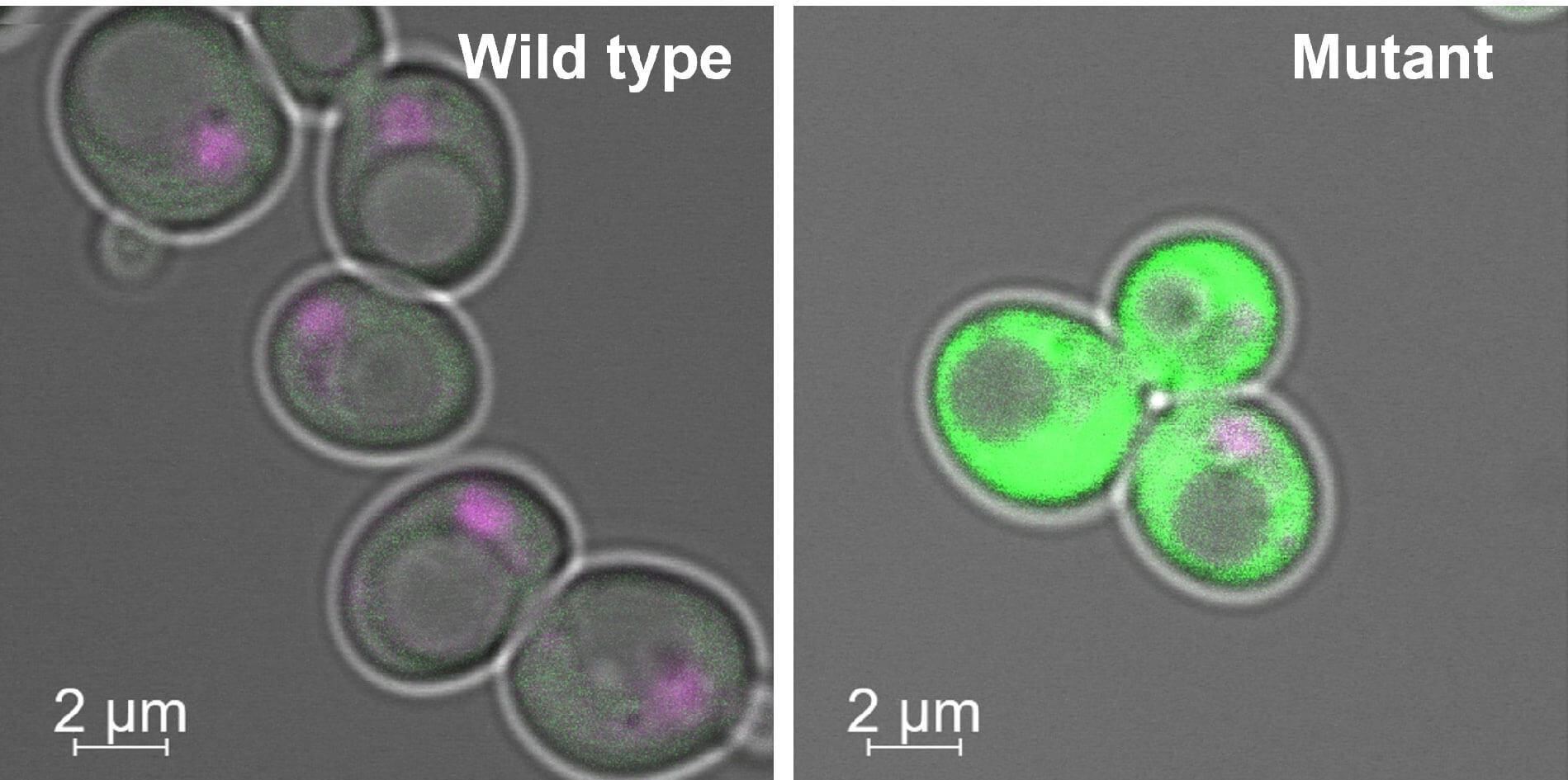
Postdocs in the Jerry Workman lab collaborate to show how chromatin “reSETs” to a silent state after a round of gene expression, blocking production of potentially harmful RNAs.
Contrary to popular lore, most scientific eureka moments don’t come in a flash to a researcher toiling alone in the lab at three in the morning. Instead, the insight required to answer a fundamental question almost always emerges from long-term collaborative effort by teams of investigators who bring varying expertise, different questions, and a lot of conversation to a problem.
Few studies illustrate the power of teamwork better than two papers from the lab of Stowers Investigator Jerry Workman, PhD, published last summer in Nature and Nature Structural and Molecular Biology. In these companion pieces, two postdocs working collaboratively described how cells protect themselves against aberrant gene expression by transcribing DNA into RNA from start to finish, rather than haphazardly initiating transcription at “cryptic” sites within a gene. These findings are noteworthy, as failure in the system could produce potentially toxic truncated RNAs, which researchers are beginning to associate with developmental defects and even cancer.
Running out of HATs
Jerry Workman has spent three decades characterizing giant protein complexes that modify the structure of chromosomes—or as scientists call it, chromatin—and control gene expression. He says the field exploded in the mid-ʼ90s as people began to characterize two classes of chromatin remodelers: the multi-subunit enzymes that activate gene expression, called histone acetylases or HATs, and their de-acetylase opponents, which repress gene expression. Workman’s lab began by concentrating on the activators.
“Initially we focused on purifying the six or seven different kinds of HAT proteins that activate gene expression in yeast,” Workman says. “At that time, every new postdoc who came to my lab got a HAT complex to characterize and then took it off to start their own lab until there were no HATs left.”
At that point, Workman started deploying new postdocs to analyze repressor complexes. By the mid-2000s studies from the lab led by former postdocs Bing Li and Mike Carozza had revealed how a yeast repressor complex called Set2 neutralized HAT activity, thereby inhibiting gene expression. Mechanistically, the Workman team showed that Set2 grabs onto the tail of the enzyme called RNA polymerase (pol II) as it moves down a DNA strand and copies it into RNA transcripts, and then shuts down gene expression in pol II’s wake, somewhat like closing the barn door after the cow passes through.
Genomically, the “door” consisted of arraying a barrier of DNA-binding proteins called histones, which when decorated with chemical acetyl groups by HATs, moved aside to allow passage of pol II, but when shorn of those groups (indirectly by Set2) blocked pol II’s access to the potentially “wrong” site on DNA.
But how the transition between acetylated versus deacetylated histones occurred at the molecular level remained unknown until the arrival of the next set of recruits to the Set2 team, namely Swami Venkatesh and Michaela (Mischa) Smolle, the authors of the 2012 studies.
Know your neighbors
Venkatesh, who led the Nature study, joined the Workman lab in 2006 to continue the studies in gene expression he began as a graduate student at Jawaharlal Nehru Centre for Advanced Scientific Research in Bangalore, India. There, Venkatesh had used human cells as a model system, but was motivated, as he says, “to move down the evolutionary ladder” to yeast after Workman visited his mentor in India and talked about Set2. Once at Stowers, Venkatesh took on the task of determining how Set2 acts to reset the histone barrier.
Smolle, the first author of the companion study, arrived a year later after completing graduate work on the structure of glucose-metabolizing enzymes at the University of Glasgow in Scotland. Unlike Venkatesh, Smolle was new to the field of chromatin biology and eager to try something new. Her first task was to conduct a proteomic screen—which she calls a “fishing expedition”—to discover what other proteins might be interacting with histones modified by Set2 in hopes of gaining insight into how those modifications occurred.
Serendipitously, Venkatesh and Smolle were assigned adjacent benches in the Workman lab, and just like lab “baymates” worldwide, they were either condemned or privileged to spend many long hours talking to each other. In their case, it was the latter, and the talk rapidly evolved from what was good in the cafeteria that day to a constant dialogue over molecular mechanisms used by Set2 to modify and shuffle histones.
“Mischa and I come from very different scientific backgrounds,” says Venkatesh. “We found it very helpful to have someone from a different field to ask about what to do. That kind of collaboration helped both of us tell a much broader story.”
Smolle agrees, noting that the Workman lab is a very team-oriented environment. “It’s not like we started out working on the same thing,” she says. “We each had our own angle on the project and our own individual expertise. In the end the two stories just merged very nicely into one.”

A new paradigm
The jumping-off point for that story was the knowledge that Set2 terminates gene expression by planting its own biochemical flag, a methyl group, on histones, which then recruits deacetylators to clip off histone acetyl groups, replace the histone barrier, and restore chromatin to a silent state, inaccessible by pol II.
From there, Venkatesh conducted a series of experiments showing that when yeast cells were made mutant in Set2 they derailed this normal gene-silencing mechanism by literally shoving “pre-acetylated” histones (sitting in reserve on the chromatin sidelines) back onto the DNA track, a stealth gene-activation move called “histone exchange.” That allowed pol II to sit wherever it liked on a DNA strand and keep gene expression going, whether it was appropriate or not.
“The old paradigm was that histones remain in place and are acetylated by HATs to allow pol II to pass and then deactylated to block it,” says Venkatesh. “But we found in a significant subset of genes the HATs weren’t required, and that histones remained acetylated as a consequence of histone exchange. That meant that one function of Set2 was to prevent histone exchange from taking place.”
The Nature study used microarray analysis to determine whether aberrations in histone acetylation were correlated with perturbed gene expression. As expected, the researchers found that unlike normal yeast, Set2 mutants produce variously truncated RNA transcripts, which could potentially gum up normal gene expression.
Meanwhile, Smolle’s proteomic “fishing expedition” reeled in a chromatin remodeler known as Isw1, which blocks the histone exchange mechanism—characterized by Venkatesh—by helping Set2 methylate histones. Strikingly, Smolle went on to show that yeast engineered to lack the Isw1 complex also exhibit perturbed gene expression marked by abnormal expression of RNA snippets rather than complete transcripts, an anomaly similar to that observed following loss of Set2.
Workman notes the studies show that the methylation mark placed on histones by Set2 works in two ways to ensure that RNA transcription does not start in the middle of a gene. “On one hand, that methylation mark recruits Isw1 to block exchange of pre-acetylated histones,” he says. “And on the other, it recruits a deacetylase to remove any acetylation marks that might happen to have sneaked in.”
The human consequences
These discoveries, made in the yeast Saccharomyces cerevisiae model system, have implications for human disease: Excessive acetylation of the histone methylated by the human Set2 counterpart occurs in several tumor types, suggesting that Set2 acts similarly to prevent recruitment of pre-acetylated histones and to ensure that pol II catalyzes RNA synthesis only where it ought to occur in mammalian cells.
“Strikingly, the human homologue of Set2, SETD2, is implicated as a tumor suppressor in breast and renal cancer, and those tumor cells are deficient in the SETD2 methylation mark,” says Workman, adding that mutations in the human homologue of Set2 are also associated with a severe developmental disorder known as Wolf-Hirschhorn syndrome. “The point is that genes that contribute to human developmental anomalies or cancer are often involved in chromosome organization. We study them in yeast to figure out what they’re doing in humans.”
Workman, who analyzes gene expression in multiple systems, says he is contemplating follow-up studies of the Set2 homologue in fruit flies. That work may, however, require a new generation of Set2 recruits, as both Smolle and Venkatesh are nearing the end of their postdoctoral training and are hunting for places to set up their own respective research labs. Venkatesh is searching for positions in the U.S., while Smolle, a native of Austria, may return to Europe.
Their collaboration has a happy outcome, but the partnering of research associates within a lab does not always end in a Hallmark moment. “It is true that highly motivated postdocs sometimes become competitors,” says Workman. “But Swami and Mischa get along well and in their case it has always been about teamwork. They spend all day together talking about their work and then come into my office, often with a great new idea I know will cost me a lot of money!”
But since 2003, when the Workman lab first reported that yeast Set2 was a histone methylase, that money has apparently been well spent. “All of these guys working on Set2 have been very interactive,” says Workman, noting that Bing Li, who now runs his own lab at University of Texas Southwestern still talks all the time with Venkatesh and Smolle. “I have been fortunate to have such a collegial group working on this project.”
News
24 June 2024
The Faculty EDGE series is just one of many advantages of the postdoctoral training program at the Stowers Institute.
Read Article
News


17 January 2024
Q&A with Stowers Postdoc Michael Church: “When I got the opportunity to join the Workman Lab, I jumped at the chance.”
Read Article
News


06 October 2023
Forty-nine members reached 20 years of service at the Stowers Institute. The Institute founders, Jim and Virginia Stowers, envisioned establishing the Institute as a long-term investment for advancing foundational knowledge in biology for the benefit of all.
Read Article